- 研究文章gydF4y2Ba
- 开放获取gydF4y2Ba
- 发表:gydF4y2Ba
huntingtin-associated蛋白质的进化40 (HAP40)与杭丁顿蛋白gydF4y2Ba
BMC进化生物学gydF4y2Ba体积gydF4y2Ba20.gydF4y2Ba文章编号:gydF4y2Ba162年gydF4y2Ba(gydF4y2Ba2020年gydF4y2Ba)gydF4y2Ba
文摘gydF4y2Ba
背景gydF4y2Ba
huntingtin-associated蛋白40 (HAP40)丰富与杭丁顿蛋白(计画),改变的蛋白质在亨廷顿氏舞蹈症(HD)。因此,我们分析的演变与计画HAP40及其交互。gydF4y2Ba
结果gydF4y2Ba
我们发现,在脊椎动物HAP40 single-exon基因编码,而在所有其他生物从multi-exon表达基因。在unikonts HAP40伴有计画,包括filastereans等gydF4y2BaCapsaspora owczarzakigydF4y2Ba和amoebozoangydF4y2Ba盘基网柄菌discoideum,gydF4y2Ba但蛋白质都缺席真菌gydF4y2Ba。gydF4y2Baunikonts以外,一些物种,例如自由amoeboflagellategydF4y2BaNaegleria gruberigydF4y2Ba,包含假定的计画和HAP40直接同源。gydF4y2Ba
我们计画显示之间的交互和生化反应HAP40延伸到鱼,和生物信息学分析进化这种交互保护提供依据。HAP40当前蛋白质数据库的最亲密的同系物是附件蛋白质可溶性N-ethylmaleimide-sensitive的家庭因素(快照)。gydF4y2Ba
结论gydF4y2Ba
我们的研究结果表明,过渡从multi-exon single-exon基因似乎发生了后移在两栖动物和脊椎动物的散度,其次是父母multi-exon基因的丧失。此外,似乎两种蛋白质可能是真核生物的根源。保护HAP40和计画及其之间的交互可能共同进化强烈表明功能这种交互的重要性。gydF4y2Ba
背景gydF4y2Ba
杭丁顿蛋白(http)是一个大型的细胞内蛋白质的分子量(MW) 348 kDa,功能涉及到不同的细胞过程。这些包括内吞作用,囊泡运输、自噬和转录调控(gydF4y2Ba1gydF4y2Ba]。的第1外显子的突变gydF4y2Ba计画gydF4y2Ba基因,结果受到多麸醯胺酸的致病性扩张呼吸道蛋白质的n端附近,导致亨廷顿氏舞蹈症(HD),一个致命的神经退行性疾病与常染色体显性遗传gydF4y2Ba2gydF4y2Ba]。gydF4y2Ba
计直接同源存在原生生物和动物,但是没有在植物和真菌gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba]。在老鼠中,计画对胚胎发育和生存能力,至关重要gydF4y2Ba计画gydF4y2Banullizygosity导致妊娠早期胚胎死亡率约为8.5天(gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba,gydF4y2Ba7gydF4y2Ba]。在斑马鱼中,gydF4y2Ba计画gydF4y2Ba功能丧失的实验导致各种表型,包括对铁代谢的影响(gydF4y2Ba8gydF4y2Ba),和不同的发展缺陷,如受损神经元的发展(gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba]。gydF4y2Ba
使用低温电子显微镜(低温电子显微镜),我们最近决定人类计画的结构(gydF4y2Ba11gydF4y2Ba],显示主要是α螺旋蛋白质与三大领域组成的串联重复序列结构图案,杭丁顿蛋白,延长因子3 (EF3),蛋白磷酸酶2 (PP2A)和酵母激酶TOR1(热)重复:n端结构域,21日热量重复(N-HEAT),一个更小的c端域12热量重复(摄氏度),和一个连接桥域。虽然同源重复组成,n端结构域形成one-and-a-half-turn超螺旋,而c端领域形成一个椭圆环。由于固有的灵活性,计画(gydF4y2Ba11gydF4y2Ba)、结构测定净化后才成为可能的计画与同源huntingtin-associated严格复杂的蛋白质40 (HAP40)。HAP40,形成4规范和2衰变tetratricopeptide重复(TPR),结合三个域之间的间隙,形成主要是疏水接触N-HEAT摄氏度的温度下和静电相互作用桥域,从而稳定构象的计画。更具体地说,结构表明,糖基HAP40包含四个带负电荷的残留物(E316、E317 E331 D333),这与一个带正电荷的区域的桥梁域。大量的计画扶少团团员(gydF4y2Ba1gydF4y2Ba,gydF4y2Ba12gydF4y2Ba计画)强烈建议作为多元互动中心协调许多不同的功能。gydF4y2Ba
在分析计画的交互和HAP40 293年人类细胞,HAP40似乎是一个非常丰富的计画[很棒gydF4y2Ba11gydF4y2Ba]。然而,我们只获得两个蛋白质之间的复杂co-expression,虽然我们无法从单独体外纯化重组蛋白(gydF4y2Ba11gydF4y2Ba]。第一次观察到的丰富,detergent-resistant交互的计画与人类细胞系HAP40彼得斯&罗斯(发表于2001年gydF4y2Ba13gydF4y2Ba]。不同寻常的交互HAP40计画,计画涉及协调所有三个领域,解释了为什么在过去HAP40很少出现大protein-interaction研究计画的很棒的,除非完整的计画是用作诱饵(gydF4y2Ba12gydF4y2Ba,gydF4y2Ba13gydF4y2Ba]。有趣的是,这两项研究之一,使用了老鼠的脑组织(gydF4y2Ba12gydF4y2Ba),发现HAP40作为计画的最丰富的关联,在少几百HTT-interacting丰富蛋白质。这些数据表明,计画的交互HAP40并不局限于人类。gydF4y2Ba
很少HAP40可用的生物功能的信息。Ras-related的一个研究小组发现这是一个效应蛋白5 (Rab5)的内吞作用,中介的Rab5-dependent招聘计画早期核内体(gydF4y2Ba14gydF4y2Ba,gydF4y2Ba15gydF4y2Ba]。gydF4y2Ba
在人类中,HAP40编码由三个sequence-identical假字因素八世intronic成绩单(gydF4y2BaF8AgydF4y2Ba)基因(gydF4y2BaF8A1gydF4y2Ba,gydF4y2BaF8A2gydF4y2Ba,gydF4y2BaF8A3gydF4y2Ba)[gydF4y2Ba16gydF4y2Ba,gydF4y2Ba17gydF4y2Ba),这都是位于X染色体中Xq28。而gydF4y2BaF8A1gydF4y2Ba假字包含在第八凝血因子的基因内区22 (gydF4y2BaF8gydF4y2Ba)基因,另两份位于外gydF4y2BaF8gydF4y2Ba基因,接近Xq端粒和间隔约495 kb和571 kb,分别gydF4y2BaF8A1gydF4y2Ba。的gydF4y2BaF8AgydF4y2Ba基因单外显子基因(凹陷),即不含有内含子,是一个更大的一部分,几乎相同的重复序列大约10 kb,名叫int22h-1 int22h-2, int22h-3。gydF4y2Ba
的gydF4y2BaF8AgydF4y2Ba基因在1990年代早期第一次描述了(gydF4y2Ba16gydF4y2Ba)时指出,内部之间的重组和基因外的副本gydF4y2BaF8AgydF4y2Ba由于导致血友病gydF4y2BaF8gydF4y2Ba基因失活。这种反演占血友病大约50%的情况下(gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba,gydF4y2Ba20.gydF4y2Ba]。gydF4y2Ba
扩展的交互HAP40计画,在净化复杂的稳定,大量的这个复杂的人类细胞培养和小鼠大脑(gydF4y2Ba12gydF4y2Ba]表明HAP40计画是一个重要的辅助因子。因为没有信息可用HAP40在不同物种的存在,我们的进化分析gydF4y2BaF8AgydF4y2Ba和其潜在的共同进化gydF4y2Ba计画gydF4y2Ba。充实功能HAP40-HTT交互的重要性,我们的研究结果强烈建议后移的参与,即染色体整合reverse-transcribed mRNA,生成的gydF4y2BaF8AgydF4y2Ba赛格在脊椎动物的进化gydF4y2BaF8AgydF4y2Ba和gydF4y2Ba计画gydF4y2Ba,保护HTT-HAP40交互。此外,我们的研究结果提供的证据同源HAP40起源和N-ethylmaleimide-sensitive因子(NSF)附件蛋白质。gydF4y2Ba
结果gydF4y2Ba
转换F8A从multi-exon single-exon基因在两栖动物和脊椎动物的散度gydF4y2Ba
而在人类和小鼠HAP40曾被证明是由单外显子基因编码(之后),有三个副本出现在人类和小鼠的只有一个,我们在研究中指出,在早期,在斑马鱼(gydF4y2Ba鲐鱼类)gydF4y2Ba的gydF4y2BaF8AgydF4y2Ba直接同源(动作:101679)包括11个外显子和宽约9200个核苷酸,而人类和老鼠中只有约1700个核苷酸。分析的出现gydF4y2BaF8AgydF4y2Ba赛格的intron-containing祖先及其可能的共存与父母的基因在某些物种,我们分析了基因组的组织和染色体定位gydF4y2BaF8AgydF4y2Ba轨迹在29代表unikonts(见附加文件gydF4y2Ba1gydF4y2Ba)。我们的分析表明,gydF4y2BaF8AgydF4y2Ba是一个赛格只有在脊椎动物,而它包含不止一个外显子在所有其他物种进行了分析。例如,一个multiple-exon组织的gydF4y2BaF8AgydF4y2Ba存在于gydF4y2Ba鲐鱼类gydF4y2Ba(11个外显子),gydF4y2Ba非洲爪蟾蜍光滑的gydF4y2Ba(12个外显子),gydF4y2BaCiona intestinalisgydF4y2Ba(18个外显子)。此外,multiple-exon组织gydF4y2BaF8AgydF4y2Ba发现在non-chordates吗gydF4y2BaAmphimedon queenslandicagydF4y2Ba(8外显子)gydF4y2Ba丝的动因gydF4y2Ba(15个外显子)。在昆虫,如gydF4y2Ba黑腹果蝇gydF4y2Ba(2外显子)gydF4y2BaBactrocera latifronsgydF4y2Ba(2外显子),一个小数量的外显子是注释。gydF4y2Ba
人类基因组变异和染色体的位置F8A直接同源gydF4y2Ba
分析完全测序的基因组拷贝数变异的gydF4y2BaF8AgydF4y2Ba在不同的物种(见附加文件gydF4y2Ba1gydF4y2Ba)。例如,在灵长类动物,人类和猩猩(gydF4y2Ba彭哥abeliigydF4y2Ba)包含三个黑猩猩(gydF4y2Ba黑猩猩gydF4y2Ba)和大猩猩(gydF4y2Ba低地大猩猩gydF4y2Ba)包含两个,和吉本斯(gydF4y2BaNomascus leucogenysgydF4y2Ba)和面容苍白的卷尾猴(gydF4y2Ba宿务capucinus模仿者gydF4y2Ba)包含一个gydF4y2BaF8AgydF4y2Ba假字。gydF4y2Ba
像家鼠(gydF4y2Ba亩骶gydF4y2Ba),只有一个gydF4y2BaF8AgydF4y2Ba直接同源检测在挪威鼠(gydF4y2Ba鼠形gydF4y2Ba)。相比之下,一个或两个gydF4y2BaF8AgydF4y2Ba在laurasiatherians直接同源存在,如猫的一个副本(gydF4y2Ba家猫silvestrigydF4y2Ba)和美洲豹(gydF4y2Ba豹属pardusgydF4y2Ba在马),一式两份(gydF4y2Ba科仕caballusgydF4y2Ba)、猪(gydF4y2Ba野猪gydF4y2Ba)、牛(gydF4y2Ba牛gydF4y2Ba)。在更多deeply-branching物种,例如在鸡(gydF4y2Ba背带吊裤带gydF4y2Ba),斑胸草雀(gydF4y2BaTaeniopygia guttatagydF4y2Ba),西方锦龟(gydF4y2BaChrysemys picta belliigydF4y2Ba),斑马鱼(gydF4y2Ba鲐鱼类gydF4y2Ba)、河豚(gydF4y2BaTakifugu摘要gydF4y2Ba),非洲爪蟾(gydF4y2Ba非洲爪蟾蜍光滑的gydF4y2Ba)和热带爪蟾(gydF4y2Ba非洲爪蟾蜍tropicalisgydF4y2Ba),我们发现只有一个直接同源。gydF4y2Ba
在简介中已经提到,3gydF4y2BaF8AgydF4y2Ba在人类的一部分重复假字(int22h-1 int22h-2, int22h-3)几乎相同的序列。另一个凹陷,H2A B1基因(组蛋白家族成员gydF4y2BaH2AFB1gydF4y2Ba),位于附近的区域gydF4y2BaF8AgydF4y2Ba。gydF4y2BaH2AFB1gydF4y2Ba编码非典型和mammalian-specific组蛋白与细胞凋亡的调控精子发生的细胞(gydF4y2Ba21gydF4y2Ba),信使rna加工、活跃转录(gydF4y2Ba22gydF4y2Ba]。当分析的基因位点gydF4y2BaF8AgydF4y2Ba直接同源,我们观察到gydF4y2BaF8AgydF4y2Ba和gydF4y2BaH2AFB1gydF4y2Ba与多个基因co-localise物种gydF4y2BaF8AgydF4y2Ba基因,如在人类、黑猩猩、猩猩、恒河猴、猪和牛。相比之下,除了猫以外,没有co-localisation的gydF4y2BaF8AgydF4y2Ba和gydF4y2BaH2AFB1gydF4y2Ba基因与一个物种gydF4y2BaF8AgydF4y2Ba基因(如长臂猿、老鼠和老鼠)。在猫,gydF4y2BaF8AgydF4y2Ba在轨迹直接同源LOC101095239周围是两个gydF4y2BaH2AFB1gydF4y2Ba假字(LOC101097798和LOC101098042),但根据我们的分析,猫拥有只有一个gydF4y2BaF8AgydF4y2Ba基因。gydF4y2Ba
在确定的染色体定位gydF4y2BaF8AgydF4y2Ba在29个代表性物种(见附加文件gydF4y2Ba1gydF4y2Ba),我们发现single-exongydF4y2BaF8AgydF4y2Ba直接同源几乎全部位于X染色体或X-chromosome-syntenic地区其他染色体。在X染色体,single-exongydF4y2BaF8AgydF4y2Ba直接同源总是靠近或在第八凝血因子的基因内区(F8)基因。在鸡,gydF4y2BaF8AgydF4y2Ba位于染色体4在2115165 - 2116487核苷酸(GRCg6a),一个轨迹已知syntenic人类吗gydF4y2BaF8AgydF4y2Ba基因和gydF4y2BaF8gydF4y2Ba轨迹。是唯一的例外,在挪威鼠(gydF4y2Ba鼠形gydF4y2Ba)gydF4y2BaF8AgydF4y2Ba和gydF4y2BaF8gydF4y2Ba都位于不同的常染色体基因染色体,即染色体1和4,分别。我们的分析表明,基因组multi-exon的本地化gydF4y2BaF8AgydF4y2Ba不是syntenic single-exon的基因组位点gydF4y2BaF8AgydF4y2Ba直接同源。例如,multi-exon的基因位点gydF4y2BaF8AgydF4y2Ba在斑马鱼是1号染色体上,而不是直接同源syntenic人类或鸡肉gydF4y2BaF8AgydF4y2Ba位点。此外,在一些物种如gydF4y2Ba鲐鱼类gydF4y2Ba,gydF4y2BaCiona intestinalis, Ciona萨维尼gydF4y2Ba,gydF4y2Ba非洲爪蟾蜍光滑的gydF4y2Ba,multi-exon的基因位点gydF4y2BaF8AgydF4y2Ba直接同源似乎non-syntenic multi-exongydF4y2BaF8AgydF4y2Ba在其他物种基因座。这种多样性基因结构、拷贝数和基因位置显示一个复杂的历史为这个家庭。gydF4y2Ba
HAP40和计画存在unikonts除了真菌gydF4y2Ba
遵循HAP40的进化和计画,我们分析了冗余蛋白质序列数据库使用PSI-BLAST或HHpred他们的存在。同意发表的数据gydF4y2Ba23gydF4y2Ba,gydF4y2Ba24gydF4y2Ba,gydF4y2Ba25gydF4y2Ba]gydF4y2Ba,gydF4y2Ba计直接同源是动物中发现的gydF4y2Ba(例如Amphimedon queenslandicagydF4y2Ba和gydF4y2Ba丝的动因gydF4y2Ba)gydF4y2Ba,gydF4y2Ba鞭虫(如。gydF4y2BaSalpingoeca罗塞塔gydF4y2Ba和gydF4y2BaMonosiga brevicollisgydF4y2Ba),filastereans(如。gydF4y2BaCapsaspora owczarzakigydF4y2Ba),ichthyosporeans(如。gydF4y2BaSphaeroforma北极蛤属gydF4y2Ba),和amoebozoans(如。gydF4y2Ba盘基网柄菌discoideumgydF4y2Ba和gydF4y2BaPlanoprotostelium fungivorumgydF4y2Ba),但不是在真菌和nucleariids(无花果。gydF4y2Ba1gydF4y2Ba和gydF4y2Ba2gydF4y2Ba额外的文件gydF4y2Ba2gydF4y2Ba,gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba)。我们认为的共同祖先unikonts包含http,蛋白质是迷失在血统导致真菌。unikonts之外,我们发现潜在的计画在一个物种的同源染色体chromalveolates(隐芽植物gydF4y2BaGuillardiaθgydF4y2Ba)和发掘(独立生存的amoeboflagellategydF4y2BaNaegleria gruberigydF4y2Ba),但不是在archaeplastidans(包括绿色植物和红藻)。鉴于潜在的分布非常不完整的计画和HAP40同系物,我们不能判断目前是否计画和HAP40真核生物的根源,在过去的真核生物的共同祖先(LECA)和植物谱系,造成的损失或起源于unikonts和横向收购少数其他血统。引人注目的是,除了parabasalidsgydF4y2Ba阴道毛滴虫gydF4y2Ba和gydF4y2BaTritrichomonas胎儿gydF4y2Ba中,我们只发现HAP40计画和HAP40相关的所有生物,完全支持的推理两种蛋白质一起进化。gydF4y2Ba

种系发生树HAP40 (gydF4y2Ba一个gydF4y2Ba)和计画(gydF4y2BabgydF4y2Ba)。HAP40或http直接同源的多重序列比对计算使用肌肉算法(gydF4y2Ba51gydF4y2Ba,gydF4y2Ba52gydF4y2Ba)在大型X软件中实现(gydF4y2Ba53gydF4y2Ba在方法部分描述。系统发育树的计算是通过贝叶斯推理MrBayes [gydF4y2Ba55gydF4y2Ba)使用杨的autocorrelatedγ模型(gydF4y2Ba56gydF4y2Ba)和混合进化模型gydF4y2Ba

改编自“生命之树Web项目”(gydF4y2Ba69年gydF4y2Ba]。分支与不确定的单系统用虚线表示gydF4y2Ba
的系统发育分布HAP40、计画和简单。在unikonts HAP40和计画主要是发现,而提前普遍存在在所有的真核生物谱系。这里显示的树gydF4y2Ba
保护模式HAP40和计画,和他们的相互作用gydF4y2Ba
调查HAP40的保护模式,计画在后生动物,我们构造的多序列比对和映射他们保护在蛋白质结构使用ConSurf服务器(gydF4y2Ba26gydF4y2Ba]。比对的HAP40 43种哺乳动物和73年non-mammals(见附加文件gydF4y2Ba7gydF4y2Ba)表示保护的N - c端区域,由一个变量脯氨酸区域(41残留在人类),它只存在于哺乳动物(图。gydF4y2Ba3gydF4y2Ba)。除此之外,这些结果反映在规范化保护与ConSurf分数计算方法(图。gydF4y2Ba4gydF4y2Ba一、表gydF4y2Ba1gydF4y2Ba额外的文件gydF4y2Ba8gydF4y2Ba,gydF4y2Ba9gydF4y2Ba,gydF4y2Ba12gydF4y2Ba)。gydF4y2Ba

多序列比对HAP40直接同源从脊椎动物代表。对齐,主要计算使用CLC工作台7,说明缺乏集中位于脯氨酸丰富地区非哺乳类物种gydF4y2Ba

进化HAP40守恒(gydF4y2Ba一个gydF4y2Ba)和计画(gydF4y2BabgydF4y2Ba):保护分数估计基于多重序列比对HAP40和http直接同源的代表物种。估计,ConSurf方法(gydF4y2Ba61年gydF4y2Ba,gydF4y2Ba62年gydF4y2Ba与贝叶斯模型[]gydF4y2Ba4gydF4y2Ba)和Jones-Taylor-Thornton模型(gydF4y2Ba70年gydF4y2Ba使用了)。只有氨基酸被郭解决et al。gydF4y2Ba11gydF4y2Ba)(蛋白质数据库标识符6 ez8)在图中所示gydF4y2Ba
在计画,N-HEAT域(残留91 - 1684)21热重复包含大量重复6和7之间插入(残留400 - 674),这是解决低温电子显微镜的结构;度高温下域(残留物,2092 - 3098年)12热量重复包含插入重复1和2之间,和重复2和3gydF4y2Ba11gydF4y2Ba]。两个域之间有桥域。ConSurf保护成绩(附加文件gydF4y2Ba10gydF4y2Ba,gydF4y2Ba11gydF4y2Ba,gydF4y2Ba12gydF4y2Ba)表明,N-HEAT插入,插入1摄氏度的温度下保存不善,插入2摄氏度的温度下和桥域显示中间保护,和N -和摄氏度的温度下的热重复最保守的蛋白质部分(无花果。gydF4y2Ba4gydF4y2Bab和表gydF4y2Ba1gydF4y2Ba)。gydF4y2Ba
在我们之前的低温电子显微镜研究中,我们指出,四个带负电荷的残留HAP40 (E316、E317 E331和D333)与积极的计画桥域(K1967, K1968, R1998、R2002 R2047) (gydF4y2Ba11gydF4y2Ba]。获得进一步的信息在这个接口,我们分析了突变的带负电荷的残留是否HAP40抑制这种交互。因此,我们计画执行下拉野生型之间的化验poly-glutamine伸展的17个谷氨酰胺(17 q-htt)和版本的HAP40,四个残留在被赖氨酸(HAP40-4 K)取代。使用HAP40(无花果。gydF4y2Ba5gydF4y2Ba)或计画(数据未显示)作为诱饵,我们的下拉交互化验证明一个缺失或强烈减少17 q-htt之间的物理相互作用和HAP40-4 K融合carboxy-terminal或伴TwinStrep-tag,分别与野生型相比HAP40(无花果。gydF4y2Ba5gydF4y2Ba)。gydF4y2Ba
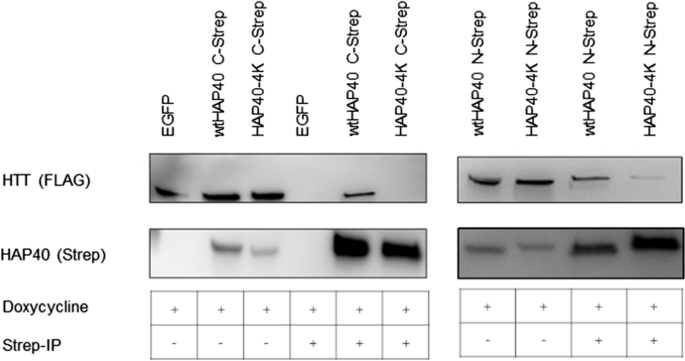
交互研究突变HAP40和计画。人类基于293 - B1.21细胞,表达17 q-htt感应,与质粒转染编码wtHAP40或HAP40突变体,E316 E317, E331,和D333被赖氨酸(HAP40-4 K)取代。在相互作用研究中,wtHAP40和HAP40融合carboxy-terminal和伴TwinStrep-tag。细胞溶解产物(IP−)或洗脱(IP +)的co-immunoprecipitation wtHAP40并使用MagStrep珠子HAP40-4 K (IBA)被免疫印迹分析。西方的屁股代表三个独立的实验gydF4y2Ba
因为这些数据进一步强调的重要性桥之间的静电作用域的计画和c端HAP40的一部分,我们通过生物信息学方法分析这些残留是否可能的进化。两个交互补丁被郭建议出版社。(gydF4y2Ba11gydF4y2Ba)之间形成一个带负电E331和D333 HAP40和带正电的R1998 R2002,和R2047计画,第二个E316与E317 HAP40和K1967 K1968计画。in-silico分析中,第一个补丁的残留显然比平均水平(表更好的保存gydF4y2Ba2gydF4y2Ba在后生动物)和主要保留收取互补,除了昆虫。相比之下,第二个补丁无法确切的残留分析由于置信区间较大,但保留收取互补性分析后生动物,除了gydF4y2BaBactrocera latifronsgydF4y2Ba,gydF4y2BaAmphimedon queenslandica,棘球绦虫multilocularisgydF4y2Ba。我们无法进一步探讨这些潜在交互基于共同进化分析(例如复杂(gydF4y2Ba27gydF4y2Ba)由于深度不足的多重比对。gydF4y2Ba
确定物理之间的相互作用在deep-branching HAP40和计画也是守恒的脊椎动物物种,我们进行下拉化验HAP40与计画gydF4y2Ba鲐鱼类gydF4y2Ba。为此,稳定HEK293-based细胞系,共同表达斑马鱼HAP40和计画,被生成。使用斑马鱼计画(无花果。gydF4y2Ba6gydF4y2Ba)或斑马鱼HAP40(数据未显示)作为诱饵,我们之间的交互分析演示了一个物理交互斑马鱼HAP40和计画。确凿HAP40-HTT交互在斑马鱼进一步的保护,我们可以co-purify斑马鱼计画与斑马鱼HAP40稳定转染HEK293TetOn细胞(数据没有显示)。gydF4y2Ba

交互研究斑马鱼HAP40和计画。斑马鱼计画与anti-FLAG磁珠免疫沉淀反应的细胞溶解产物DrHTT-HAP40细胞。DrHTT-HAP40细胞持续表达的斑马鱼HAP40计画和斑马鱼的表达可以通过添加强力霉素诱导。细胞溶解产物(IP−),洗出液(IP +)分析了通过免疫印迹分析使用anti-Strep (HAP40)或anti-FLAG (http)抗体。西方的屁股代表三个独立的实验gydF4y2Ba
综上所述,我们的研究表明,生物信息学分析和生化作用的物理相互作用HAP40计画是进化保守至少在脊椎动物,但可能在动物和,基于严格的同现的两种蛋白质,还煞有介事地在unikonts。gydF4y2Ba
HAP40和NSF附件蛋白质同源gydF4y2Ba
自HAP40生物功能的信息是有限的,我们寻找它的同源染色体在已知结构的蛋白质,利用隐马尔可夫模型。搜索与HAP40从人类被播种,斑马鱼和果蝇。最好的比赛,用概率值> 99%,附件是哺乳动物N-ethylmaleimide-sensitive因素蛋白质α(SNAPA)和γ(SNAPG),和他们的酵母直接同源Sec17。吸附蛋白质调节囊泡瞄准和融合通过编排提前受体蛋白之间的相互作用(陷阱)和胞质蛋白N-ethylmaleimide-sensitive因素(NSF) (gydF4y2Ba28gydF4y2Ba,gydF4y2Ba29日gydF4y2Ba,gydF4y2Ba30.gydF4y2Ba]。的临时家庭是在真核生物中广泛存在,许多物种组成多个假字;例如,酵母包含一个同族体(Sec17),人类包含三个同系物(SNAPA、SNAPB SNAPG) (gydF4y2Ba31日gydF4y2Ba]。gydF4y2Ba
除了代表最好的打击HAP40序列空间,快照也其最佳冲击结构比较(图。gydF4y2Ba7gydF4y2Ba)。最好的匹配,搜索HAP40-like结构RCSB蛋白质数据银行使用大理网络服务器(gydF4y2Ba32gydF4y2Ba),SNAPA, SNAPG, Sec17, z得分在16和18之间,和均方根偏差(rmsd) 2.5和3之间。相比较而言,下一个最好的比赛是相当糟糕,从z得分13和5 A的表示。像HAP40,拍摄由6 TPR发夹但缺乏插入重复2和脯氨酸地区发现哺乳动物HAP40重复4和5之间。gydF4y2Ba

HAP40和吸附蛋白质的结构。答:结构对齐HAP40(蓝色,6 ez8 PDB标识符,gydF4y2Ba11gydF4y2Ba与Sec17])(红色,PDB标识符1 qqe, (gydF4y2Ba71年gydF4y2Ba]),SNAPA直接同源gydF4y2Ba酿酒酵母gydF4y2BaHAP40, B:表面电荷分布,SNAPA, SNAPG从不同角度的红色着色表示一个负电荷和蓝色表示积极的表面电荷。说明三维结构的生成与嵌合体1.13.1 [X版本gydF4y2Ba63年gydF4y2Ba]gydF4y2Ba
这些比较表明,HAP40拍对方的近亲在蛋白质数据库中。这种同源关系可能导致两个家庭有一个共同的祖先与6 TPR发夹或从他们的独立放大相似结构从一个祖先,单一的TPR发夹。我们已经讨论了这两个场景之前(gydF4y2Ba33gydF4y2Ba]。全球TPR发夹的序列相似性gydF4y2BangydF4y2Ba一个家庭的匹配最接近TPR发夹gydF4y2Ban 'gydF4y2Ba其他的指示前的场景,而序列相似性的一个家庭的所有TPR发夹匹配彼此更紧密地比其他家庭的任何重复表明后者。重复的详细比较内部和之间的两个家庭(无花果。gydF4y2Ba8gydF4y2Ba)表明,就有一个明确的信号放大从单个TPR重复以来发夹gydF4y2BangydF4y2Ba匹配概率高的蛋白质内的其他重复。HAP40,没有相应的内部比赛,每个重复匹配只有本身有高概率。相反,在self-match之外,每个HAP40重复都有其最佳匹配的等效重复。这表明,上述两种场景发生在快照的进化和HAP40:初始放大了家庭,这强烈差异化的一种新形式的一个分支(HAP40),因此,其祖先在一个完全像拉断TPR蛋白形成的。gydF4y2Ba

成对嗯比较HAP40和TPR发夹。四个规范人类HAP40 TPR发夹,发夹1,3,4,6,相应的发夹酵母吸附家族的蛋白质Sec17使用HHpred相互比较gydF4y2Ba
根据观察到的序列和结构相似性,我们调查了三个人类是否吸附蛋白质SNAPA SNAPB, SNAPG与人类的计画。从下拉化验HEK293TetOn细胞细胞溶解产物,共同表达17 q-htt SNAPA一起SNAPB,或SNAPG,我们不能发现任何交互使用17 q-htt(无花果。gydF4y2Ba9gydF4y2Ba)或吸附的蛋白质作为诱饵(数据没有显示)。没有检测到的交互并不奇怪,因为快照显示不同表面电荷分布比HAP40(无花果。gydF4y2Ba7gydF4y2Bab)。gydF4y2Ba
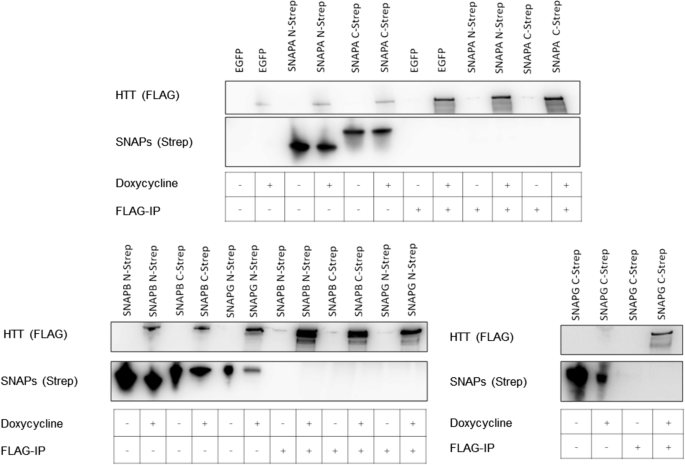
相互作用的研究计画与SNAPA、SNAPB或SNAPG使用17 q-htt作为诱饵。计画B1.21细胞诱导表达和基于pBSK-CMV的质粒转染表达与羧基吸附,伴TwinStrep-Tags。在转染后48 h快照co-immunoprecipitated使用国旗珠子和免疫印迹分析。西方的屁股代表三个独立的实验gydF4y2Ba
讨论gydF4y2Ba
在人类基因组中,之后,占大约8.9%的蛋白编码基因(gydF4y2Ba34gydF4y2Ba]。HAP40、赛格在人类表达从三个sequence-identical single-exon假字位于X染色体,一个在和两个邻居的gydF4y2BaF8gydF4y2Ba基因。我们的基因组组织的分析gydF4y2BaF8AgydF4y2Ba基因在真核生物表明只有脊椎动物拥有轨迹gydF4y2BaF8AgydF4y2Ba基因是一个凹陷,这表明转换从multi-exon基因(MEG) single-exon基因(凹陷)发生期间或之后的散度两栖动物和脊椎动物。gydF4y2Ba
不同机制的出现凹陷multi-exon前体讨论了。据一位,之后出现重复事件intron-containing基因(gydF4y2Ba35gydF4y2Ba),主要是单外显子复制通常导致截断(gydF4y2Ba36gydF4y2Ba]。情况并非如此gydF4y2BaF8AgydF4y2Ba由于凹陷和梅格直接同源代码同源蛋白质几乎相同的大小。另一种理论假设之间的同源重组之后,出现反向转录mRNA的中间值和相应的基因的基因位点(gydF4y2Ba37gydF4y2Ba]。我们的发现gydF4y2BaF8AgydF4y2Ba基因位点single-exon multi-exon配置并不是syntenic反对这样一个机制,虽然不能排除次要易位事件。后移的reverse-transcribed mRNA已被建议作为主要的生成机制功能活跃之后,从父母multi-exon前兆,导致retrogenes(也叫retrocopies) [gydF4y2Ba34gydF4y2Ba,gydF4y2Ba38gydF4y2Ba]。intron-loss和观察到的基因位点之间缺乏同线性赛格在脊椎动物和梅格gydF4y2BaF8AgydF4y2Ba基因在斑马鱼和其他non-amniotic物种表明后移的intron-less基因拷贝到一个新的轨迹发生,而原来的梅格gydF4y2BaF8AgydF4y2Ba轨迹迷路了。缺乏多聚腺苷酸序列不反对这种机制,自后移在早期非哺乳类物种,如鸡(gydF4y2Ba39gydF4y2Ba),似乎是由逆转录病毒机制而不是长点缀元素会导致集成的多聚腺苷酸序列(gydF4y2Ba38gydF4y2Ba]。gydF4y2Ba
引人注目的是,灵长类动物和一些laurasiatherians拥有不止一个gydF4y2BaF8AgydF4y2Ba假字,人类是包含在一个更大的int22h重复。基本上排除这样一种可能性,即蛋白质3假字不同的表达功能,身份的三个序列gydF4y2BaF8AgydF4y2Ba假字在人类是由基因转换gydF4y2Ba19gydF4y2Ba),即假字之间的非互惠性的遗传物质转移由于同源性。大多数中性基因的复制有一个低概率成为固定在一个人口和经常丢失或进化功能不活跃的假基因(gydF4y2Ba40gydF4y2Ba]。复制gydF4y2BaF8AgydF4y2Ba基因可能是保留在进化过程中由于重复事件可能赋予更高的进化人类灵长类动物和laurasiatherians健身。然而,没有数据支持的固定gydF4y2BaF8AgydF4y2Ba复制报告。这是可以想象的gydF4y2BaF8AgydF4y2Ba基因可以增加蛋白质表达水平(gydF4y2Ba41gydF4y2Ba]或subfunctionalisation [gydF4y2Ba42gydF4y2Ba,gydF4y2Ba43gydF4y2Ba]带来进化的优势;16 s rrna [gydF4y2Ba41gydF4y2Ba和组蛋白基因gydF4y2Ba40gydF4y2Ba)例子授予进化优势的高表达水平由于基因的复制。另一方面,subfunctionalisation被假设为固定高度相似的假字的原因(gydF4y2Ba40gydF4y2Ba,gydF4y2Ba43gydF4y2Ba]。例如,不同的假字可能获得不同的组织,development-specific表达谱如图所示engrailed-1和engrailed-1b [gydF4y2Ba40gydF4y2Ba,gydF4y2Ba42gydF4y2Ba假字。自gydF4y2BaF8A1gydF4y2Ba基因位于内含子的22gydF4y2BaF8gydF4y2BaF8转录基因反义方向,HAP40表达水平可能受转录活性的影响gydF4y2BaF8gydF4y2Ba基因,基因在肝细胞被强烈的表达。然而,作为放大和固定的另一种解释gydF4y2BaF8AgydF4y2Ba在灵长类动物和一些laurasiatherians假字,也有可能是相当紧密的联系gydF4y2BaH2AFB1gydF4y2Ba组蛋白基因可能授予一个进化优势,因为它一直在观察其他组蛋白基因(gydF4y2Ba40gydF4y2Ba]。gydF4y2Ba
在蛋白质水平,人类HAP40包含一个内部脯氨酸地区的41个氨基酸长度,根据我们的数据,在non-mammals缺席。在哺乳动物中,这一地区是非常变量长度和氨基酸组成。这个地区由低温电子显微镜一直悬而未决,指示的灵活性,和是可有可无的交互HAP40计画,由co-expression如图所示,在人类细胞中蛋白质相互作用研究(gydF4y2Ba11gydF4y2Ba]。经常脯氨酸图案直接参与蛋白质-蛋白质之间的关系(gydF4y2Ba44gydF4y2Ba)和交互的识别蛋白质绑定到本地区重大利益解开mammalian-specific HAP40的函数。gydF4y2Ba
HAP40身体与计画在细胞显著水平,如图所示为人类细胞体外培养(gydF4y2Ba13gydF4y2Ba),和体内小鼠大脑gydF4y2Ba12gydF4y2Ba]。在这项研究中,我们显示的物理相互作用斑马鱼HAP40和计画,当人类HEK293TetOn细胞中表达。我们建议这种交互进化是守恒的,基于接口的分析在我们的低温电子显微镜观察到的结构。几个带电氨基酸预测研究调解之间的交互的c端地区HAP40(负电荷)和桥梁领域的计画在许多直接同源(正电荷)是守恒的,和他们的突变HAP40减少计画的交互。gydF4y2Ba
当提出HAP40 RAB5的效应,其生物功能的信息在很大程度上仍然是难以捉摸的。最近确定低温电子显微镜HTT-HAP40复杂结构及其热行为展开相比单独个体蛋白质(gydF4y2Ba11gydF4y2Ba]表明HAP40起结构作用,协调计画的三个领域,屏蔽大暴露疏水表面区域分布在一个大型计画的一部分。自进化关系可以用来推断假设蛋白功能,我们旨在识别HAP40同源染色体与已知函数,这可能为HAP40提供额外的功能。在我们的分析中,发现HAP40拍TPR-containing蛋白质的相似性最高的序列和结构层次(无花果。gydF4y2Ba7gydF4y2Ba),提高的可能性HAP40也可以参与膜泡运输。符合这个推理,Pal等人提出的数据表明HAP40介导的招聘计画和Ras-related蛋白5 (RAB5)早期核内体gydF4y2Ba14gydF4y2Ba,gydF4y2Ba15gydF4y2Ba]。尽管如此,因为TPR-containing蛋白质表现出高度多样化的函数作为脚手架蛋白(gydF4y2Ba45gydF4y2Ba,gydF4y2Ba46gydF4y2Ba),目前还不清楚到什么程度的势函数HAP40可以推断其同源性快照,快照以来更不能取代HAP40在其与计画(本研究)。gydF4y2Ba
结论gydF4y2Ba
这项研究是第一个分析HAP40的进化,其编码基因因素八世intronic转录基因(gydF4y2BaF8AgydF4y2Ba)和潜在的共同进化HAP40计画。HAP40由single-exon编码基因(凹陷)在脊椎动物,而从multi-exon基因表达(MEG)在所有其他生物。在unikonts HAP40伴有计画,包括filastereans等gydF4y2BaCapsaspora owczarzakigydF4y2Ba和amoebozoangydF4y2Ba盘基网柄菌discoideum,gydF4y2Ba但蛋白质都缺席真菌gydF4y2Ba。gydF4y2Baunikonts以外,一些物种,例如自由amoeboflagellategydF4y2BaNaegleria gruberigydF4y2Ba,包含假定的计画和HAP40直接同源,提高两种蛋白质进化的可能性在真核生物的根源。gydF4y2Ba
计和HAP40之间的交互,这是显示在人类和老鼠,还延伸到鱼,和生物信息学分析这种交互进化的保护提供依据。HAP40当前蛋白质数据库的最亲密的相同器官是附件蛋白质可溶性N-ethylmaleimide-sensitive的家庭因素(快照)。断了,然而,无法取代HAP40与计画的交互。综上所述,保护HAP40和计画及其之间的交互可能共同进化强烈显示功能的这种交互的重要性。gydF4y2Ba
方法gydF4y2Ba
识别F8A和http直接同源不同的分类群gydF4y2Ba
确定直接同源的gydF4y2BaF8AgydF4y2Ba和gydF4y2Ba计画gydF4y2Ba在不同的物种代表真核生物的进化,人类HAP40序列比较(NP_036283.2)和计画(NP_002102)参考序列是由不连续的megablast PSI-BLAST [gydF4y2Ba47gydF4y2Ba),或HHpred [gydF4y2Ba48gydF4y2Ba,gydF4y2Ba49gydF4y2Ba]。对nr70_euk10Jun PSI-BLAST,执行搜索(2019)数据库使用人类HAP40 (UniProt ID: P23610)查询序列,BLOSUM60矩阵,1 e−3和一个创造价值的价值包含1 e−3的阈值。在MPI HHpred搜索进行了生物信息学工具针对PDB_mmCIF70数据库使用默认设置,除了目标序列的数量:1000。此外,我们执行搜索HAP40和http直接同源,与人类的序列被确定初始搜索或者在NCBI的蛋白质数据库,数据库搜索gydF4y2Ba果蝇,Amphimedon queenslandicagydF4y2Ba,gydF4y2BaNaegleria gruberigydF4y2Ba。我们确认所有的确认达到基于他们的整体序列相似性,序列长度,和产生的e-values搜索BALST或HHpred,和全球两两序列比对的目视检查相应的查询序列使用Needleman-Wunsch算法实现的欧洲生物信息学研究所(EMBL-EBI) [gydF4y2Ba50gydF4y2Ba]。gydF4y2Ba
系统发育树的计算gydF4y2Ba
蛋白质序列HAP40和计画(额外的文件gydF4y2Ba2gydF4y2Ba)与肌肉算法(gydF4y2Ba51gydF4y2Ba,gydF4y2Ba52gydF4y2Ba10.0.5]使用大型X软件(版本,构建# 10180924 - x86_64) (gydF4y2Ba53gydF4y2Ba]。肌肉的算法,公开处罚−2.9和差距扩大处罚(0)使用。此外,疏水性的乘数为1.20,未加权的paired-group意味着16迭代算法(UPGMA)和最大和最小使用对角线的长度24gydF4y2Ba51gydF4y2Ba,gydF4y2Ba52gydF4y2Ba,gydF4y2Ba54gydF4y2Ba]。系统模型的计算是通过贝叶斯推理实现3.2.7 MrBayes版本。(gydF4y2Ba55gydF4y2Ba]。推理,杨的autocorrelatedγ模型(gydF4y2Ba56gydF4y2Ba使用)和混合进化模型。分析了1000000 (http)或500000 (HAP40)代在巴登-符腾堡州的高性能计算集群(BwHPC)。系统发育树的融合是由分裂频率的标准差衡量树的两个独立样本之间的相似性。HAP40标准差的分裂频率计画的0.011880和0.001884分裂频率的标准偏差。此外,为系统推理的可靠性测试,我们计算的系统发育树未加权的两组方法与算术平均值算法的最大吝啬法(UPGMA)和实现大型X的所有软件(gydF4y2Ba53gydF4y2Ba(数据未显示)。gydF4y2Ba
分析F8A基因结构及其基因位点gydF4y2Ba
HAP40基因结构分析使用国家生物技术信息中心(NCBI)的基因组浏览器。染色体的位置和外显子的数量评估在所有选定代表性物种(附加文件gydF4y2Ba1gydF4y2Ba),被选出的代表unikonts的主要类群。如果gydF4y2BaF8AgydF4y2Ba基因位于x染色体上,,gydF4y2BaF8AgydF4y2Ba基因位点进行分析,同线性使用比较基因组学工具提供的运用(97年版)(gydF4y2Ba57gydF4y2Ba]。自gydF4y2BaH2AFTB1gydF4y2Ba位于int22-h在人类和重复吗gydF4y2BaH2AFTB1gydF4y2Ba直接同源只是描述了在哺乳动物中,我们评估的本地化gydF4y2BaF8AgydF4y2Ba相对于gydF4y2BaH2AFTB1gydF4y2Ba在所有选定的哺乳动物。gydF4y2Ba
在一些物种中(gydF4y2Ba秀丽隐杆线虫,Amphimedon queenslandica, Bactrocera latrifrons Ciona intestinalis,果蝇,棘球绦虫multilocularis, Exaiptasia pallida,日本血吸虫,gydF4y2Ba和gydF4y2Ba丝的动因gydF4y2Ba),gydF4y2BaF8AgydF4y2Baorthologues被确定蛋白质序列,但不是在核苷酸水平通过不连续的megablast使用信使rna序列gydF4y2Ba智人gydF4y2Ba或gydF4y2Ba非洲爪蟾蜍tropicalisgydF4y2Ba。因为一些gydF4y2BaF8AgydF4y2Ba可能不带注释的副本,我们排除了这些物种的分析基因副本的数量,以避免不完整的基因注释结果的影响。gydF4y2Ba
存在直接同源HAP40脯中部地区的不同gydF4y2Ba
脯氨酸中部地区的评估存在HAP40直接同源代表性物种,在蛋白质数据库中搜索NCBI与蛋白质进行基本的局部比对搜索工具(爆炸)对宏基因组蛋白质提供的NCBI数据库。人类HAP40的氨基酸序列(加入数字:NP_036283.2)被用作查询序列。进行深入的分析,主要的哺乳动物和non-mammals分类群,我们分析了HAP40序列物种属于常见的哺乳动物和类non-mammals订单。因此,序列的43种哺乳动物和73 non-mammals(见附加文件gydF4y2Ba7gydF4y2Ba)被当地两两序列比对分析均为算法对人类参考序列(gydF4y2Ba58gydF4y2Ba)使用两两序列比对工具的欧洲生物信息学研究所(gydF4y2Ba59gydF4y2Ba)和50块替换矩阵(gydF4y2Ba60gydF4y2Ba]。central-proline富裕地区的存在是由手工检查的检查结果比对。gydF4y2Ba
进化与ConSurf保护方法的估计gydF4y2Ba
保护ConSurf分数计算的方法(gydF4y2Ba26gydF4y2Ba,gydF4y2Ba61年gydF4y2Ba,gydF4y2Ba62年gydF4y2Ba)基于多个代表HAP40或者http直接同源序列比对(见附加文件gydF4y2Ba8gydF4y2Ba,gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba和gydF4y2Ba11gydF4y2Ba肌肉)计算的算法部分的解释“计算系统发育树”。保护分数归一化标准差的1和0的意思是[gydF4y2Ba61年gydF4y2Ba]。消极保护的分数表明更高的进化保护相比其他残留的蛋白质。计算保护分数投射到HTT-HAP40蛋白质结构被郭et al。gydF4y2Ba11gydF4y2Ba6 ez8)] (PDB数据库标识符。分子图形和分析与加州大学旧金山分校嵌合体(1.13.1版),由生物运算资源,开发可视化、信息学加州大学,旧金山,在国家卫生研究院的支持下P41-GM103311 [gydF4y2Ba63年gydF4y2Ba]。gydF4y2Ba
分析保护分数使用R执行脚本语言(gydF4y2Ba64年gydF4y2Ba)和r程序ggplot2 [gydF4y2Ba65年gydF4y2Ba]和readr [gydF4y2Ba66年gydF4y2Ba]。算术均值和标准的错误保护得分的均值特定氨基酸残基的蛋白质域计算(附加文件gydF4y2Ba12gydF4y2Ba)。计分析域的位置被从郭et al。gydF4y2Ba11gydF4y2Ba]。gydF4y2Ba
HMM-based比较HAP40和TPR发夹gydF4y2Ba
调查的进化起源HAP40和临时的家庭gydF4y2Ba,gydF4y2Ba我们评估序列相似性的TPR发夹。我们选择了四个规范人类HAP40 TPR发夹,发夹1,3,4,6,相应的发夹在酵母吸附蛋白质家族Sec17代表。我们第一次搜查了nr70数据库(NCBI冗余蛋白质序列数据库集群在70%序列身份)人类HAP40和酵母的同系物Sec17使用爆炸[gydF4y2Ba47gydF4y2Ba,gydF4y2Ba67年gydF4y2Ba),价值阈值(安勤科技)设置为0.001和对齐覆盖率(-qcov_hsp_perc) 50%。单独的多序列比对HAP40和Sec17同系物是解析获得的点击率。这两个阵营随后被用于提取个人比对TPR发夹。资料隐马尔可夫模型(摘要)计算使用hhmake从联盟并与hhsearch(二级结构得分是关闭),既HH-suite3软件包的敏感序列搜索基于摘要(gydF4y2Ba49gydF4y2Ba]。gydF4y2Ba
互动研究人类HAP40和17 qhtt突变gydF4y2Ba
B1.21 HEK293TetOn-based细胞系(gydF4y2Ba68年gydF4y2Ba)是诱导72 h 1µg /毫升强力霉素和转染,使用聚乙烯亚胺,与基于pBSK-CMV质粒编码wtHAP40 HAP40-4 K (p。[Glu316Lys; Glu317Lys Glu331Lys;Asp333Lys])与羧基-或伴TwinStrep-tag融合。控制,B1.21细胞转染质粒编码EGFP。细胞收集DPBS离心10分钟400×紧随其后gydF4y2BaggydF4y2Ba。消息灵通的细胞颗粒细胞溶解25毫米,300毫米氯化钠,渐变20 0.5%,蛋白酶抑制剂(罗氏),pH值8.0 30分钟。溶菌产物是通过离心20000×10分钟gydF4y2BaggydF4y2Ba在4°C或3 h和孵化MagStrep珠子(IBA)或磁anti-Flag珠子(西格玛奥德里奇)。珠子与25毫米洗了三次玫瑰,300毫米氯化钠,0.02%渐变20,pH值8.0。结合蛋白与预筛选了SDS加载缓冲区。样本分析使用anti-Flag与sds - page及免疫印迹分析(西格玛奥德里奇,1:2500稀释)和anti-Strep抗体(IBA, 1:1000稀释)。gydF4y2Ba
代的稳定细胞系co-expression计画和HAP40斑马鱼gydF4y2Ba
cDNA,人类codon-optimized斑马鱼和编码长度计画(NCBI NP_571093)和c端融合到他的亲和力标签,是由DNA合成(Thermofisher)。的cDNA克隆到质粒载体pTRE-Tight-BI-AcGFP1 (Clontech)允许co-expression zebrafish-HTT和GFP感应与强力霉素(阿霉素)。结果分析和瞬态表达质粒被限制在293个细胞。HEK293细胞Tet-ON (Clontech) co-transfected线性化表达质粒和线性化选择潮霉素抗性基因的质粒编码。阳性细胞克隆被潮霉素选择孤立。单克隆细胞株表达zebrafish-HTT (drHTT)通过有限稀释法,验证了免疫印迹分析细胞溶解产物使用单克隆抗体anti-FLAG(西格玛奥德里奇)。gydF4y2Ba
表达质粒pBSK / CMV-drHAP40TS建于表达启动子血巨细胞病毒的控制,人类codon-optimized斑马鱼HAP40 (NCBI XP_005160094.1) c端Twin-Strep-tag融合。一起drHTT细胞转染pBSK / CMV-drHAP40TS嘌呤霉素抗性的质粒编码。Puromycin-resistant细胞克隆被孤立和单克隆细胞株(drHTT-HAP40 3-2-26)导出了有限稀释。drHTT-HAP40 3-2-26细胞被免疫印迹分析验证细胞溶解产物与单克隆anti-FLAG ab(西格玛奥德里奇,1:2500稀释)检测Dox-inducible drHTT表达式,和一个anti-Strep抗体(Iba, 1:1000)检测本构drHAP40表达式。gydF4y2Ba
互动研究HAP40和http直接同源鲐鱼类gydF4y2Ba
zHTT-zHAP40 HEK293-based细胞系的诱导与1µg /毫升强力霉素72 h。不诱导zHTT-HAP40细胞被用作控制。下拉化验进行描述。gydF4y2Ba
互动研究SNAPA SNAPB SNAPG和人类的野生型计画gydF4y2Ba
前所述HEK293TetOn-based B1.21细胞,表达17 qhtt感应与强力霉素(gydF4y2Ba68年gydF4y2Ba)与pBSK-CMV-based转染质粒表达人类SNAPA SNAPB或SNAPG羧基-或伴融合Twin-Strep标签。下拉化验使用国旗珠像之前描述的那样进行。gydF4y2Ba
可用性的数据和材料gydF4y2Ba
所有分析核苷酸和蛋白质序列可以从核苷酸或蛋白质数据库检索的国家生物技术信息中心(NCBI)加入数字给出额外的文件。此外,比对用于计算序列的系统发育树和保护被添加到额外的文件。gydF4y2Ba
缩写gydF4y2Ba
- F8:gydF4y2Ba
-
凝血因子八世gydF4y2Ba
- F8A:gydF4y2Ba
-
VIII因子intronic成绩单gydF4y2Ba
- 绿色荧光蛋白:gydF4y2Ba
-
绿色荧光蛋白gydF4y2Ba
- H2AFB1:gydF4y2Ba
-
H2A组蛋白家族成员B1基因gydF4y2Ba
- HAP40:gydF4y2Ba
-
Huntingtin-associated蛋白质40gydF4y2Ba
- 高清:gydF4y2Ba
-
亨廷顿氏舞蹈症gydF4y2Ba
- 热:gydF4y2Ba
-
杭丁顿蛋白,延长因子3,脂类和蛋白磷酸酶2 a激酶TORgydF4y2Ba
- HHpred:gydF4y2Ba
-
不对比较同源性检测和结构预测gydF4y2Ba
- 计画:gydF4y2Ba
-
杭丁顿蛋白gydF4y2Ba
- LECA:gydF4y2Ba
-
最后的真核生物的共同祖先gydF4y2Ba
- 梅格:gydF4y2Ba
-
Multi-exon基因gydF4y2Ba
- NSF:gydF4y2Ba
-
N-ethylmaleimide-sensitive因素gydF4y2Ba
- PSI-BLAST:gydF4y2Ba
-
Position-specific基本迭代局部搜索定位工具gydF4y2Ba
- RAB5:gydF4y2Ba
-
Ras相关蛋白质5gydF4y2Ba
- 赛格:gydF4y2Ba
-
Single-exon基因gydF4y2Ba
- SNAPA:gydF4y2Ba
-
可溶性N-ethylmaleimide-sensitive蛋白α因子依恋gydF4y2Ba
- SNAPB:gydF4y2Ba
-
可溶性N-ethylmaleimide-sensitive附件蛋白β因素gydF4y2Ba
- SNAPG:gydF4y2Ba
-
附件蛋白γ可溶性N-ethylmaleimide-sensitive因素gydF4y2Ba
- 快照:gydF4y2Ba
-
附件蛋白质可溶性N-ethylmaleimide-sensitive因素gydF4y2Ba
- TPR:gydF4y2Ba
-
Tetratricopeptide-like螺旋域gydF4y2Ba
引用gydF4y2Ba
Saudou F,亨伯特杭丁顿蛋白的生物学。神经元。2016;89:910-26。gydF4y2Bahttps://doi.org/10.1016/j.neuron.2016.02.003gydF4y2Ba。gydF4y2Ba
亨廷顿氏舞蹈症合作研究小组。小说包含的三核苷酸重复基因扩展和亨廷顿氏舞蹈症染色体不稳定。细胞。1993;72:971 - 83。gydF4y2Bahttps://doi.org/10.1016/0092 - 8674 (93) 90585 - egydF4y2Ba。gydF4y2Ba
Tartari M, Gissi C罗Sardo V,祖C, Picardi E, Pesole G,均大肠系统发育比较杭丁顿蛋白同系物揭示了一种原始的外观polyQ海胆。另一个星球杂志。2008;25:330-8。gydF4y2Bahttps://doi.org/10.1093/molbev/msm258gydF4y2Ba。gydF4y2Ba
Mayrose我Graur D Ben-Tal N, Pupko t比较特定站点rate-inference蛋白质序列的方法:经验贝叶斯方法优越。另一个星球杂志。2004;21:1781 - 91。gydF4y2Bahttps://doi.org/10.1093/molbev/msh194gydF4y2Ba。gydF4y2Ba
小'Kusky阿纳西尔J, Floresco某人Diewert VM,理查德JM, Zeisler J, et al。亨廷顿氏舞蹈症基因的靶向破坏导致胚胎杀伤力和杂合的行为和形态变化。细胞。1995;81:811-23。gydF4y2Bahttps://doi.org/10.1016/0092 - 8674 (95) 90542 - 1gydF4y2Ba。gydF4y2Ba
Duyao MP,奥尔巴赫AB,瑞安,Persichetti F,巴恩斯GT,麦克尼尔公司SM, et al。鼠标亨廷顿氏舞蹈症基因的失活Hdh同系物。科学。1995;269:407-10。gydF4y2Bahttps://doi.org/10.1126/science.7618107gydF4y2Ba。gydF4y2Ba
蔡特林,刘JP,查普曼DL Papaioannou VE、Efstratiadis a细胞凋亡增加,小鼠早期胚胎杀伤力nullizygous亨廷顿氏舞蹈症基因同系物。Nat麝猫。1995;11:155 - 63。gydF4y2Bahttps://doi.org/10.1038/ng1095 - 155gydF4y2Ba。gydF4y2Ba
拉姆斯登,Henshall TL,达扬年代,Lardelli MT,理查兹国际扶轮。Huntingtin-deficient斑马鱼展览缺陷铁利用和发展。哼摩尔麝猫。2007;16:1905-20。gydF4y2Bahttps://doi.org/10.1093/hmg/ddm138gydF4y2Ba。gydF4y2Ba
Henshall TL,塔克B,拉姆斯登,布拉克年代,Lardelli MT,理查兹国际扶轮。选择性神经元发展中斑马鱼杭丁顿蛋白的要求。哼摩尔麝猫。2009;18:4830-42。gydF4y2Bahttps://doi.org/10.1093/hmg/ddp455gydF4y2Ba。gydF4y2Ba
阿克曼H, Anichtchik,弗莱明富特M,戈德史密斯P,罗奇,Rubinsztein博士。BDNF水平下降是一个主要贡献者杭丁顿蛋白击倒斑马鱼的胚胎表型。J > 2009; 29:1343-9。gydF4y2Bahttps://doi.org/10.1523/jneurosci.6039 - 08.2009gydF4y2Ba。gydF4y2Ba
郭问,本·H,程J,来自M,断T, Pfeifer G, et al .杭丁顿蛋白的低温电子显微镜结构。大自然。2018;555:117-20。gydF4y2Bahttps://doi.org/10.1038/nature25502gydF4y2Ba。gydF4y2Ba
格林尼Shirasaki DI,呃,Al-Ramahi我灰色的M, Boontheung P, Geschwind DH, et al。网络组织在哺乳动物大脑杭丁顿蛋白的蛋白质组interactome。神经元。2012;75:41-57。gydF4y2Bahttps://doi.org/10.1016/j.neuron.2012.05.024gydF4y2Ba。gydF4y2Ba
彼得斯MF,罗斯CA。隔离40-kDa Huntingtin-associated蛋白质。生物化学杂志。2001;276:3188 - 94。gydF4y2Bahttps://doi.org/10.1074/jbc.M008099200gydF4y2Ba。gydF4y2Ba
朋友,雪华铃F,效果,Zerial m .监管核内体动力学Rab5和Huntingtin-HAP40效应在生理和病理条件下复杂。方法Enzymol。2008; 438:239-57。gydF4y2Bahttps://doi.org/10.1016/s0076 - 6879 (07) 38017 - 8gydF4y2Ba。gydF4y2Ba
朋友,雪华铃F, lom B,舍瓦,Zerial m . Huntingtin-HAP40复杂是一种新型Rab5效应,调节早期内体能动性和亨廷顿氏舞蹈症的上调。J细胞杂志。2006;172:605-18。gydF4y2Bahttps://doi.org/10.1083/jcb.200509091gydF4y2Ba。gydF4y2Ba
莱文森B, Kenwrick年代,Lakich D,哈蒙德G, Gitschier j .转录基因内含子的人为因素八世基因。基因组学。1990;7:1-11。gydF4y2Bahttps://doi.org/10.1016/0888 - 7543 (90) 90512 - sgydF4y2Ba。gydF4y2Ba
莱文森B, Kenwrick年代,选定P,费舍尔K, Gitschier j .第三个证据记录从人类VIII因子基因。基因组学。1992;14:585-9。gydF4y2Bahttps://doi.org/10.1016/s0888 - 7543 (05) 80155 - 7gydF4y2Ba。gydF4y2Ba
Naylor J, Brinke,草丛,绿点,Giannelli f .特点mRNA异常中发现严重的血友病患者的一半是由于大的反演。哼摩尔麝猫。1993;2:1773-8。gydF4y2Ba
Bagnall RD、艾尔斯KL、绿点,Giannelli f基因转换和演化中Xq28 duplicons参与循环倒置造成严重血友病a基因组研究》2005;15:214-23。gydF4y2Bahttps://doi.org/10.1101/gr.2946205gydF4y2Ba。gydF4y2Ba
鼻咽粘膜de CD,鲍文DJ。分子的特征基因内区22第八凝血因子基因的同源染色体:一个更新。J Thromb Haemost。2008; 6:1822-4。gydF4y2Bahttps://doi.org/10.1111/j.1538-7836.2008.03094.xgydF4y2Ba。gydF4y2Ba
顾C,通问,郑L,梁Z, Pu J,梅H, et al . TSEG-1,小说的成员组蛋白H2A变异,通过促进精子发生的细胞凋亡参与精子发生。基因组学。2010;95:278 - 89。gydF4y2Bahttps://doi.org/10.1016/j.ygeno.2010.02.005gydF4y2Ba。gydF4y2Ba
Tolstorukov我,高盛JA,吉尔伯特C, PJ Ogryzko V,金斯敦再保险、公园。H2A组蛋白变体。bdd与活跃在人类细胞中转录和mRNA加工。摩尔细胞。2012;47:596 - 607。gydF4y2Bahttps://doi.org/10.1016/j.molcel.2012.06.011gydF4y2Ba。gydF4y2Ba
Gissi C, Pesole G,均E, Tartari m .杭丁顿蛋白的基因进化脊索动物及其独特的特性在海鞘类Ciona属。BMC基因组学。2006;7:288。gydF4y2Bahttps://doi.org/10.1186/1471 - 2164 - 7 - 288gydF4y2Ba。gydF4y2Ba
Myre马,拉姆斯登,汤普森MN, Wasco W,麦克唐纳我,Gusella摩根富林明。杭丁顿蛋白的缺乏社会变形虫Dictyostelium discoideum多效性的影响。公共科学图书馆麝猫。2011;7:e1002052。gydF4y2Bahttps://doi.org/10.1371/journal.pgen.1002052gydF4y2Ba。gydF4y2Ba
祖C,均大肠正常的杭丁顿蛋白的功能。琼斯:贝茨克,Tabrizi年代,L,编辑。亨廷顿氏舞蹈症。第四。牛津大学,纽约:牛津大学出版社;2014年。p . 243 - 73。gydF4y2Ba
Ashkenazy H, Abadi年代,Martz E,伞形花耳草啊,Mayrose我Pupko T, Ben-Tal n ConSurf 2016:一种改进的方法来估计和可视化进化在大分子保护。核酸研究》2016;44:W344-50。gydF4y2Bahttps://doi.org/10.1093/nar/gkw408gydF4y2Ba。gydF4y2Ba
霍普夫助教,绿色AG,舒伯特B, Mersmann年代,Scharfe CPI,英格JB, et al . EVcouplings Python框架共同进化的序列分析。生物信息学。2019;35:1582-4。gydF4y2Bahttps://doi.org/10.1093/bioinformatics/bty862gydF4y2Ba。gydF4y2Ba
Kaiser CA, Schekman r .截然不同的交会基因控制运输囊泡形成和融合早期分泌途径。细胞。1990;61:723-33。gydF4y2Bahttps://doi.org/10.1016/0092 - 8674 (90) 90483 - ugydF4y2Ba。gydF4y2Ba
女孩IC, Schekman R,罗斯曼我,凯撒CA。酵母SEC17基因产物功能相当于哺乳动物alpha-SNAP蛋白质。生物化学杂志。1992;267:12106-15。gydF4y2Ba
Sollner T Whiteheart SW Brunner M, Erdjument-Bromage H, Geromanos年代,Tempst P, Rothman我。提前受体参与囊泡瞄准和融合。大自然。1993;362:318-24。gydF4y2Bahttps://doi.org/10.1038/362318a0gydF4y2Ba。gydF4y2Ba
鼠尾草属的植物做什么,女孩IC,罗斯曼我。断了,一个家庭的NSF附件蛋白参与细胞内的膜融合在动物和酵母。细胞。1990;61:709-21。gydF4y2Bahttps://doi.org/10.1016/0092 - 8674 (90) 90482 - tgydF4y2Ba。gydF4y2Ba
河中沙洲l .基准折DaliLite 5卷的检测。生物信息学》2019。gydF4y2Bahttps://doi.org/10.1093/bioinformatics/btz536gydF4y2Ba。gydF4y2Ba
朱H,赛普维达E,哈特曼医学博士Kogenaru M, Ursinus, Sulz E, et al。重复折叠蛋白质的起源本质上从一个无序的祖先。Elife》2016。gydF4y2Bahttps://doi.org/10.7554/eLife.16761gydF4y2Ba。gydF4y2Ba
Jorquera R, R奥尔蒂斯,Ossandon F, Cardenas JP,赛普维达R,冈萨雷斯C,福尔摩斯DS。SinEx DB:针对单一外显子编码序列在哺乳动物基因组数据库。数据库(牛津大学)。2016年。gydF4y2Bahttps://doi.org/10.1093/database/baw095gydF4y2Ba。gydF4y2Ba
张你们Vibranovski医学博士克里斯凯BH, m . retrocopy的警示标识:intron-containing基因的dna复制大大有助于单外显子基因的起源。生物信息学。2011;27:1749-53。gydF4y2Bahttps://doi.org/10.1093/bioinformatics/btr280gydF4y2Ba。gydF4y2Ba
爱默生JJ, Kaessmann H, Betran E,长m .广泛的哺乳动物X染色体上基因的流量。科学。2004;303:537-40。gydF4y2Bahttps://doi.org/10.1126/science.1090042gydF4y2Ba。gydF4y2Ba
吉尔伯特·w·罗伊·西南的进化spliceosomal内含子:模式、谜题和进步。Nat牧师麝猫。2006;7:211-21。gydF4y2Bahttps://doi.org/10.1038/nrg1807gydF4y2Ba。gydF4y2Ba
Kaessmann H, Vinckenbosch N,长m . RNA-based基因重复:机械和进化的见解。Nat牧师麝猫。2009;10:19-31。gydF4y2Bahttps://doi.org/10.1038/nrg2487gydF4y2Ba。gydF4y2Ba
国际鸡基因组测序协作组。鸡基因组的序列和比较分析提供了独特的视角在脊椎动物进化。大自然。2004;432:695 - 716。gydF4y2Bahttps://doi.org/10.1038/nature03154gydF4y2Ba。gydF4y2Ba
张静重复基因的进化:一个更新。趋势生态另一个星球。2003;18:292-8。gydF4y2Bahttps://doi.org/10.1016/s0169 - 5347 (03) 00033 - 8gydF4y2Ba。gydF4y2Ba
Klappenbach是的,邓巴JM,施密特TM。核糖体rna操纵子拷贝数反映了细菌的生态策略。:环境Microbiol。2000; 66:1328-33。gydF4y2Bahttps://doi.org/10.1128/aem.66.4.1328 - 1333.2000gydF4y2Ba。gydF4y2Ba
力,林奇M,皮科特FB,爱慕,燕YL, Postlethwait j .保存重复基因的互补,退行性突变。遗传学。1999;151:1531-45。gydF4y2Ba
林奇M,迫使a重复基因的概率由subfunctionalization保存。遗传学。2000;154:459 - 73。gydF4y2Ba
威廉姆森MP。脯氨酸地区蛋白质的结构和功能。j . 1994; 297 (Pt 2): 249 - 60。gydF4y2Bahttps://doi.org/10.1042/bj2970249gydF4y2Ba。gydF4y2Ba
D 'Andrea LD,里根l . TPR蛋白:通用的螺旋。生物化学科学。2003;趋势28:655 - 62。gydF4y2Bahttps://doi.org/10.1016/j.tibs.2003.10.007gydF4y2Ba。gydF4y2Ba
Perez-Riba Itzhaki LS。tetratricopeptide-repeat图案是一个通用的平台,使不同的分子识别模式。当今结构生物学观点》2019;54:43-9。gydF4y2Bahttps://doi.org/10.1016/j.sbi.2018.12.004gydF4y2Ba。gydF4y2Ba
张Altschul科幻,马登TL,谢弗AA, J,张Z,米勒W, Lipman DJ。有缺口的爆炸和PSI-BLAST:新一代的蛋白质数据库搜索程序。核酸研究》1997;25:3389 - 402。gydF4y2Ba
齐默尔曼L,斯蒂芬斯,南S-Z劳D, Kubler J,尹浩然,Lozajic。一个完全重新实现MPI生物信息学工具和一个新的HHpred服务器的核心。J杂志。2018;430:2237-43。gydF4y2Bahttps://doi.org/10.1016/j.jmb.2017.12.007gydF4y2Ba。gydF4y2Ba
Steinegger米、迈耶米,米尔迪塔族人M, Vohringer H, Haunsberger SJ, sod j . HH-suite3快速远程同源性检测和蛋白质注释。BMC Bioinform。2019; 20:473。gydF4y2Bahttps://doi.org/10.1186/s12859 - 019 - 3019 - 7gydF4y2Ba。gydF4y2Ba
Chojnacki年代,考利,李J, Foix,洛佩兹r编程访问生物信息学工具EMBL-EBI更新:2017。核酸研究》2017;45:W550-3。gydF4y2Bahttps://doi.org/10.1093/nar/gkx273gydF4y2Ba。gydF4y2Ba
埃德加钢筋混凝土。肌肉:多重序列比对方法和降低时间和空间复杂度。BMC生物信息学。2004;5:113。gydF4y2Bahttps://doi.org/10.1186/1471 - 2105 - 5 - 113gydF4y2Ba。gydF4y2Ba
埃德加钢筋混凝土。肌肉:多重序列比对和高准确性和高吞吐量。核酸研究》2004 b; 32:1792-7。gydF4y2Bahttps://doi.org/10.1093/nar/gkh340gydF4y2Ba。gydF4y2Ba
Kumar年代,Stecher G,李米,Knyaz C,田村k .大型X:分子进化遗传学分析计算平台。另一个星球杂志。2018;35:1547-9。gydF4y2Bahttps://doi.org/10.1093/molbev/msy096gydF4y2Ba。gydF4y2Ba
华G-J,挂个氯、林bxcy吴fc,陈Y-W,唐CY。MGUPGMA:快速与多个图形处理单元使用NCCL UPGMA算法。另一个星球Bioinform网上。2017;13:1176934317734220。gydF4y2Bahttps://doi.org/10.1177/1176934317734220gydF4y2Ba。gydF4y2Ba
Huelsenbeck JP Ronquist f . MRBAYES:贝叶斯推理的系统发育树。生物信息学。2001;17:754-5。gydF4y2Bahttps://doi.org/10.1093/bioinformatics/17.8.754gydF4y2Ba。gydF4y2Ba
杨z的时空过程模型的DNA序列的进化。遗传学。1995;139:993 - 1005。gydF4y2Ba
Achuthan Zerbino博士,P, Akanni W, Amode先生,巴雷尔D,扎拉拉难民营J, et al . 2018年运用。核酸研究》2018;46:d754 - 61。gydF4y2Bahttps://doi.org/10.1093/nar/gkx1098gydF4y2Ba。gydF4y2Ba
史密斯TF,沃特曼女士共同分子子序列的识别。J杂志。1981;147:195-7。gydF4y2Bahttps://doi.org/10.1016/0022 - 2836 (81) 90087 - 5gydF4y2Ba。gydF4y2Ba
李坎迪娅·迈克威廉H, W, Uludag M, Squizzato年代,公园YM,足丝N, et al。分析工具从EMBL-EBI Web服务。核酸研究》2013;41:w597 - 600。gydF4y2Bahttps://doi.org/10.1093/nar/gkt376gydF4y2Ba。gydF4y2Ba
Henikoff年代,Henikoff詹。从蛋白质氨基酸替换矩阵块。《美国国家科学院刊。1992;89:10915-9。gydF4y2Bahttps://doi.org/10.1073/pnas.89.22.10915gydF4y2Ba。gydF4y2Ba
格拉泽F, Pupko T,巴斯,贝尔再保险,Bechor-Shental D, Martz E, Ben-Tal n ConSurf:识别蛋白质的功能区域地表测绘系统发育信息。生物信息学。2003;19:163-4。gydF4y2Bahttps://doi.org/10.1093/bioinformatics/19.1.163gydF4y2Ba。gydF4y2Ba
朗道M, Mayrose我,罗森博格Y,格拉泽F, Martz E, Pupko T, Ben-Tal n ConSurf 2005:进化的投影保护大量的残留蛋白质结构。核酸研究》2005;33:w299 - 302。gydF4y2Bahttps://doi.org/10.1093/nar/gki370gydF4y2Ba。gydF4y2Ba
EF佩特森工作室内由手工制作完成,戈达德TD,黄CC,沙发GS,格林布拉特DM,孟EC, Ferrin TE。加州大学旧金山分校Chimera-a可视化系统进行探索性的研究和分析。J第一版杂志。2004;25:1605-12。gydF4y2Bahttps://doi.org/10.1002/jcc.20084gydF4y2Ba。gydF4y2Ba
R核心团队。接待员:统计计算的语言和环境。奥地利维也纳;2020年。gydF4y2Ba
韦翰h . ggplot2:优雅的图形进行数据分析。可汗:施普林格;2016年。gydF4y2Ba
韦翰H,海丝特J,弗朗索瓦·r·readr:阅读矩形文本数据;2018年。gydF4y2Ba
马卡马乔C, Coulouris G, Avagyan V, N,帕帕多普洛斯J,比尔K,马登TL。爆炸+:体系结构和应用程序。BMC生物信息学。2009;10:421。gydF4y2Bahttps://doi.org/10.1186/1471 - 2105 - 10 - 421gydF4y2Ba。gydF4y2Ba
董黄B,卢卡斯T, Kueppers C、X, Krause M, Bepperling,等。可伸缩的生产在人类细胞和生化特征的完整的正常和突变杭丁顿蛋白。PLoS ONE。2015年。gydF4y2Bahttps://doi.org/10.1371/journal.pone.0121055gydF4y2Ba。gydF4y2Ba
麦迪森博士,舒尔茨钴。生命之树Web项目。2007年。gydF4y2Bahttps://www.tolweb.orggydF4y2Ba。gydF4y2Ba
琼斯DT,泰勒WR,桑顿JM。从蛋白质序列突变数据矩阵的快速生成。第一版:Biosci。1992; 8:275 - 82。gydF4y2Ba
大米LM共舞。蛋白质晶体结构的膜泡运输Sec17:影响吸附功能网罗复杂的拆卸。摩尔细胞。1999;4:85 - 95。gydF4y2Ba
确认gydF4y2Ba
这项工作进行的计算资源bwUniCluster由铁道部科学研究和艺术和大学的巴登-符腾堡州,德国,BwHPC框架内程序。我们感谢Steffen只是为了讨论斑马鱼HAP40 Axel Furstberger,约翰·克劳斯和汉斯Kestler首次生物信息学工作。gydF4y2Ba
资金gydF4y2Ba
开放获取资金启用并由Projekt交易。这项工作由德意志Forschungsgemeinschaft (DFG,德国研究基金会——项目编号412854449)和马克斯·普朗克的社会。Q.G.,W.B. and R.F.-B. have received funding from the European Commission (FP7 GA ERC-2012-SyG_318987-ToPAG). R.F.-B. acknowledges funding from the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) through Germany’s Excellence Strategy—EXC 2067/1- 390729940. The funding agencies provided research support and did not influence study design, collection, analysis, and interpretation of data and writing of the manuscript.
作者信息gydF4y2Ba
作者和联系gydF4y2Ba
贡献gydF4y2Ba
弗吉尼亚州和SK设计生化实验女士和女士,艾尔,SK计划in-silico分析。TE和黑洞生成HEK293-based细胞系DrHTT DrHTT-HAP40。女士和TE co-immunoprecipitation实验用斑马鱼进行计画和斑马鱼HAP40。In-silico分析(两两序列比对,系统发育分析,分析的蛋白质与ConSurf保护,和分析f8a基因及其基因位点)是由硕士结构对齐Sec17 HAP40是由女士,路上和SK Co-Immunoprecipitation实验与SNAPA SNAPB, SNAPG和17 q-htt由女士和TE和HAP40-4 K女士进行的实验,VA HAP40 HMM-based进行比较和TPR发夹。in-silico和生化实验的数据分析是由女士,弗吉尼亚州,艾尔和SK、女士和VA准备数据。女士,弗吉尼亚州,艾尔和SK手稿和女士写道,弗吉尼亚州,黑洞,TE,世行,路上,RF-B,艾尔,SK修订手稿和评论。所有作者阅读和批准最终的手稿。gydF4y2Ba
相应的作者gydF4y2Ba
道德声明gydF4y2Ba
伦理批准和同意参与gydF4y2Ba
不适用。gydF4y2Ba
同意发布gydF4y2Ba
不适用。gydF4y2Ba
相互竞争的利益gydF4y2Ba
作者宣称没有利益冲突。gydF4y2Ba
额外的信息gydF4y2Ba
出版商的注意gydF4y2Ba
施普林格自然保持中立在发表关于司法主权地图和所属机构。gydF4y2Ba
补充信息gydF4y2Ba
额外的文件1。gydF4y2Ba
在不同的物种f8a直接同源。基因被不连续的megablast使用f8a从智人(NM_012151.3)或非洲爪蟾蜍tropicalis (NM_001078703.1)。gydF4y2Ba
额外的文件2。gydF4y2Ba
加入的数量确定HAP40和http直接同源。gydF4y2Ba
额外的文件3。gydF4y2Ba
对齐的HAP40直接同源用于系统发育分析。gydF4y2Ba
额外的文件4。gydF4y2Ba
系统发育树HAP40直接同源。gydF4y2Ba
额外的文件5。gydF4y2Ba
对齐的计画直接同源用于系统发育分析。gydF4y2Ba
额外的文件6。gydF4y2Ba
系统发育树的计画直接同源。gydF4y2Ba
额外的文件7。gydF4y2Ba
物种用于分析中央HAP40脯地区的存在。蛋白质序列HAP40直接同源众议员哺乳动物和非哺乳类物种被当地pairwise-sequence比对分析。gydF4y2Ba
额外的文件8。gydF4y2Ba
对齐的HAP40直接同源用于ConSurf分析。gydF4y2Ba
额外的文件9。gydF4y2Ba
生为HAP40 ConSurf分析的结果。gydF4y2Ba
额外的文件10。gydF4y2Ba
对齐的计画直接同源用于ConSurf分析。gydF4y2Ba
额外的文件11。gydF4y2Ba
原计画ConSurf分析的结果。gydF4y2Ba
额外的文件12。gydF4y2Ba
R脚本用于计算领域的保护。gydF4y2Ba
权利和权限gydF4y2Ba
开放获取gydF4y2Ba本文是基于知识共享署名4.0国际许可,允许使用、共享、适应、分布和繁殖在任何媒介或格式,只要你给予适当的信贷原始作者(年代)和来源,提供一个链接到创作共用许可证,并指出如果变化。本文中的图片或其他第三方材料都包含在本文的创作共用许可证,除非另有说明在一个信用额度的材料。如果材料不包括在本文的创作共用许可证和用途是不允许按法定规定或超过允许的使用,您将需要获得直接从版权所有者的许可。查看本许可证的副本,访问gydF4y2Bahttp://creativecommons.org/licenses/by/4.0/gydF4y2Ba。知识共享公共领域奉献豁免(gydF4y2Bahttp://creativecommons.org/publicdomain/zero/1.0/gydF4y2Ba)适用于数据可用在这篇文章中,除非另有说明在信贷额度的数据。gydF4y2Ba
关于这篇文章gydF4y2Ba

引用这篇文章gydF4y2Ba
来自,M。阿尔瓦,V。,Huang, B.et al。gydF4y2Bahuntingtin-associated蛋白质的进化40 (HAP40)与杭丁顿蛋白。gydF4y2BaBMC另一个星球杂志gydF4y2Ba20.gydF4y2Ba162 (2020)。https://doi.org/10.1186/s12862 - 020 - 01705 - 5gydF4y2Ba
收到了gydF4y2Ba:gydF4y2Ba
接受gydF4y2Ba:gydF4y2Ba
发表gydF4y2Ba:gydF4y2Ba
DOIgydF4y2Ba:gydF4y2Bahttps://doi.org/10.1186/s12862 - 020 - 01705 - 5gydF4y2Ba
关键字gydF4y2Ba
- 杭丁顿蛋白gydF4y2Ba
- Huntingtin-associated蛋白质40gydF4y2Ba
- 附件蛋白质可溶性N-ethylmaleimide-sensitive因素gydF4y2Ba
- 后移gydF4y2Ba
- Single-exon基因gydF4y2Ba
- 分子进化gydF4y2Ba
- 蛋白质共同进化gydF4y2Ba